Fighting against human trafficking
The writer is a social activist who has been at the forefront of the fight against human trafficking and has led NGOs that have helped rescue hundreds of victims across India.

The writer is a social activist who has been at the forefront of the fight against human trafficking and has led NGOs that have helped rescue hundreds of victims across India.

Software companies operate in the private sector, with profit as a main motive. Yet, any organisation is expected to follow certain ethical practices with its employees in the implementation of which its HR team plays an important role.

Whether it was to be a doctor by day or a homicide detective by night, I was painting pictures far beyond my ability and, not to mention reality. When I ultimately couldn't decide, I chose to do the most heroic thing. I took up science in class 11.

The writer's commitment to her art is evident from the outset with her choice of subject and racy style. The author has demonstrated intelligence in choosing a pertinent topic that one can relate to in today’s fast-paced, tech-driven city life.

Cinematic endeavours wield the power to transport audiences across epochs, offering glimpses into foregone eras and shedding light on the history of human experience.

Speak of Segu outside Segu, but do not speak of Segu in Segu.

Well, April has descended on us, and the words return to haunt. Today I ponder the cruelty that we have let loose. Cruelty of different hues. Of the worst kind is the cruelty we have subjected nature to

Rousseau writes in The Social Contract, “The social pact, far from destroying natural equality, substitutes, on the contrary, a moral and lawful equality for whatever physical inequality that nature may have imposed on mankind; so that however unequal in strength and intelligence, men become equal by covenant and by right.”
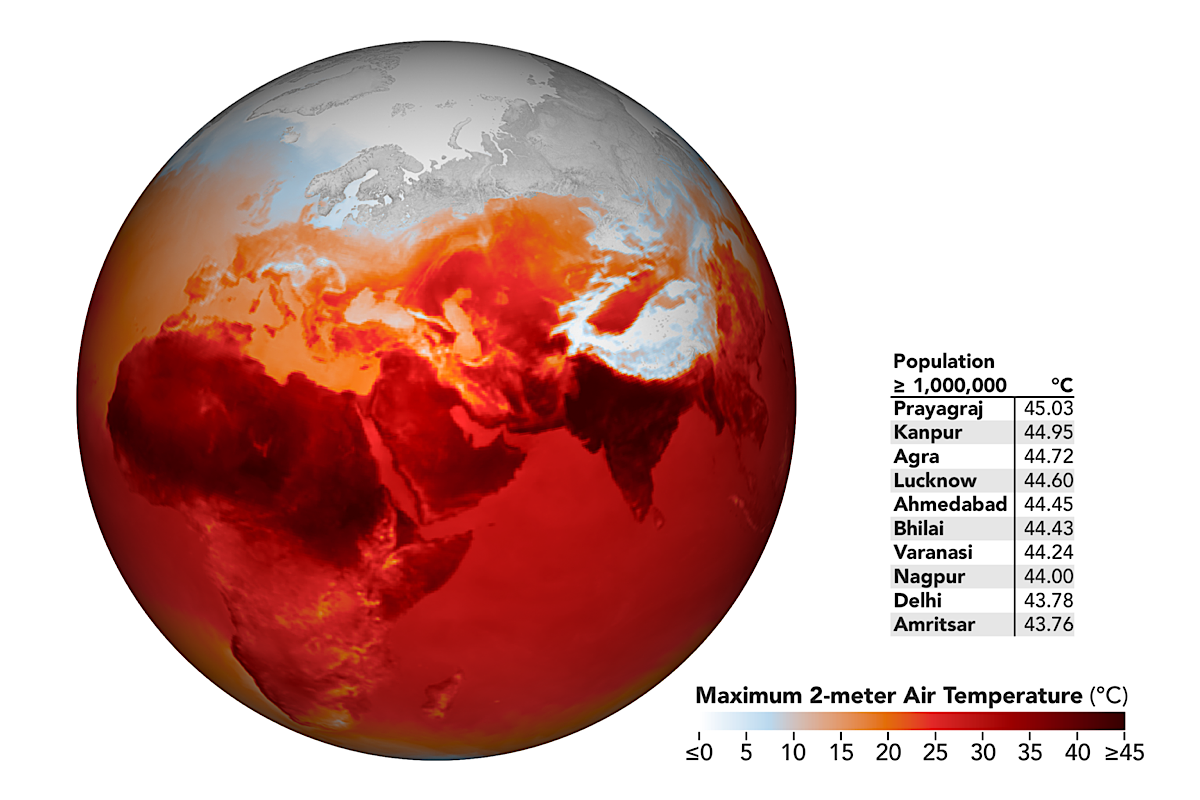
On the day of the incident, the closest Met stations recorded a maximum temperature of 34–38 °C, along with a relatively high humidity (45 per cent), exacerbating the impact of the heat.

The year 2023 was the hottest year on record, with a global mean temperature of 1.45 °C above pre-industrial values. Due to El Niño conditions in 2023, all days were more than 1ºC above the pre-industrial level




