
Microscopic life: From chaos to classification
Micro-organisms constitute a very antique group of living organisms which appeared on the Earth's surface almost 3000 million years ago.
Heres how the three enzymes called RNA polymerases I, II, and III carry out transcription in the eukaryotic nucleus.

Some of the properties of the three RNA polymerases that function in the nucleus of the eukaryotic cell, as well as two others found in mitochondria and chloroplasts. The nuclear enzymes are designated RNA polymerases I, II, and III. As the table indicates, these enzymes differ in their location within the nucleus and in the kinds of RNA they synthesise.
The nuclear RNA polymerases also differ in their sensitivity to various inhibitors, such as -amanitin, a deadly toxin produced by the mushroom Amanita phalloides (known commonly as the “death cap” or “destroying angel”).
Advertisement
RNA polymerase I resides in the nucleolus and is responsible for synthesising an RNA molecule that serves as a precursor for three of the four types of rRNA found in eukaryotic ribosomes (28S rRNA, 18S rRNA, and 5.8S rRNA). This enzyme is not sensitive to -amanitin. Its association with the nucleolus is understandable, as the nucleolus is the site of ribosomal RNA synthesis and ribosomal subunit assembly.
Advertisement
A prominent structural component of the eukaryotic nucleus is the nucleolus, the ribosome factory of the cell. Typical eukaryotic cells contain one or two nucleoli, but the occurrence of several more is not uncommon. The nucleolus is usually a spherical structure measuring several micrometers in diameter, but wide variations in size and shape are observed.
Because of their relatively large size, nucleoli are easily seen with the light microscope and were first observed more than 200 years ago. However, it was not until the advent of electron microscopy in the 1950s that the structural components of the nucleolus were clearly identified. In thin-section electron micrographs, each nucleolus appears as a membrane-free organelle consisting of fibrils and granules. The fibrils contain DNA that is being transcribed into ribosomal RNA (rRNA), the RNA component of ribosomes.
The granules are rRNA molecules being packaged with proteins (imported from the cytoplasm) to form ribosomal subunits. As we saw earlier, the ribosomal subunits are subsequently exported through the nuclear pores to the cytoplasm. Because of their role in synthesising RNA, nucleoli become heavily radiolabeled when the cell is exposed to radioactive precursors of RNA.
RNA polymerase II is found in the nucleoplasm and synthesises precursors to mRNA, the class of RNA molecules that code for proteins. In addition, RNA polymerase II synthesises most of the snRNAs-small nuclear RNAs involved in posttranscriptional RNA processing. Thus, polymerase II is responsible for the synthesis of the greatest variety of RNA molecules. The enzyme is extremely sensitive to – amanitin, which explains the toxicity of this compound to humans and other animals.
RNA polymerase III is also a nucleoplasmic enzyme, but it synthesises a variety of small RNAs, including tRNA precursors and the smallest type of ribosomal RNA, 5S rRNA. Mammalian RNA polymerase III is sensitive to a-amanitin, but only at higher levels of the toxin than are required to inhibit RNA polymerase II. (The comparable enzymes of some other eukaryotes, such as insects and yeasts, are insensitive to -amanitin.)
Structurally, RNA polymerases I, II, and III are somewhat similar to each other as well as to prokaryotic core RNA polymerase. The three enzymes are all quite large, with multiple polypeptide subunits and molecular weights around 500,000. RNA polymerase II, for example, has more than ten subunits of at least eight different types.
The three biggest subunits are evolutionarily related to the prokaryotic RNA polymerase subunits,? and / ?’. Three of the smaller subunits lack that relationship but are also found in RNA polymerases II and III. The RNA polymerases of mitochondria and chloroplasts resemble their prokaryotic counterparts closely, as you might expect from the probable origins of these organelles as endosym-biotic bacteria. Like bacterial RNA polymerase, they are resistant to -amanitin.
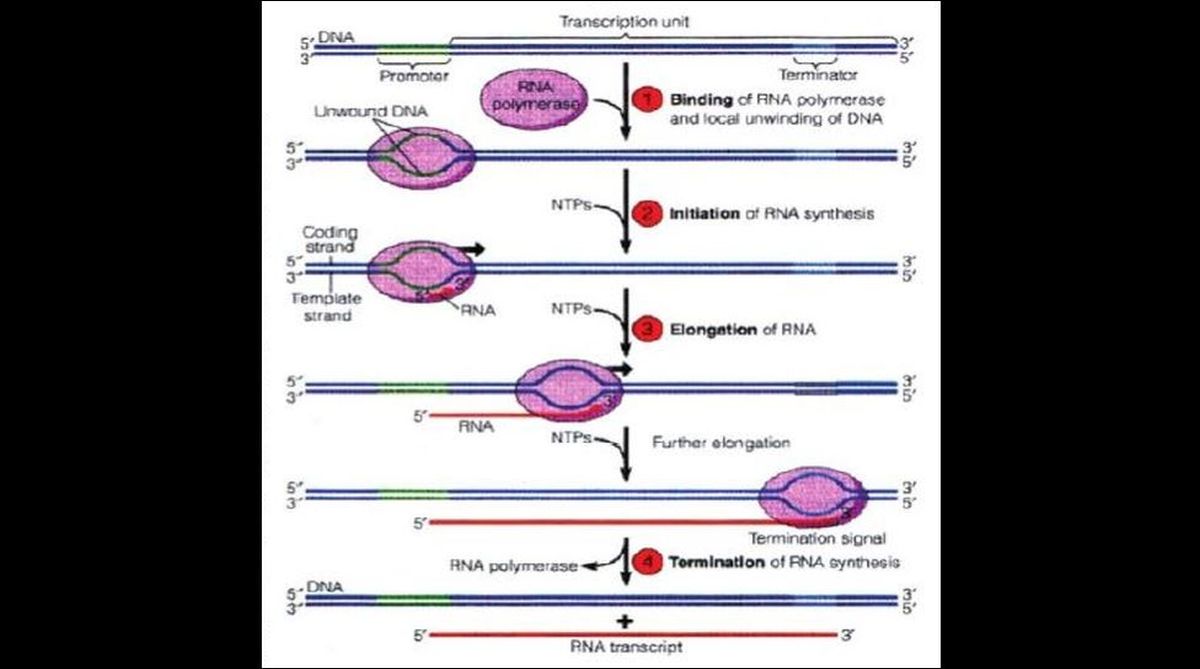
Transcription in eukaryotic cells involves the same four stages but the process in eukary-otes is more complicated than that in prokaryotes. The main differences are as follows:
The writer is associate professor and head, department of botany, Ananda Mohan College.
Advertisement












