India’s Gita Sabharwal appointed UN Resident Coordinator in Indonesia
India's Gita Sabharwal has been appointed as the United Nations Resident Coordinator in Indonesia, with the host Government's approval.

India's Gita Sabharwal has been appointed as the United Nations Resident Coordinator in Indonesia, with the host Government's approval.

If you have spent nights eating chocolates or ice cream, then 'loneliness' can be the reason behind the binging on sugary items, say researchers.

This annual event serves as a crucial platform for facilitating collaboration and innovation in this rapidly evolving field.

The 17-year-old Gukesh is just one win away from winning this year's tournament. He defeated the French GM in Round 13.

Tech firm Logitech India's head of B2B Anand Lakshmanan on Sunday said that the adoption of hybrid work models offers a significant opportunity for the country's video conferencing market.

Gaming app platforms like Dream11 and My 11 Circle have shown an impressive surge in both daily active cash users and revenue per person.

His comments come amid recent attacks on Israel by Iran, reportedly in retaliation to the attack on their embassy.
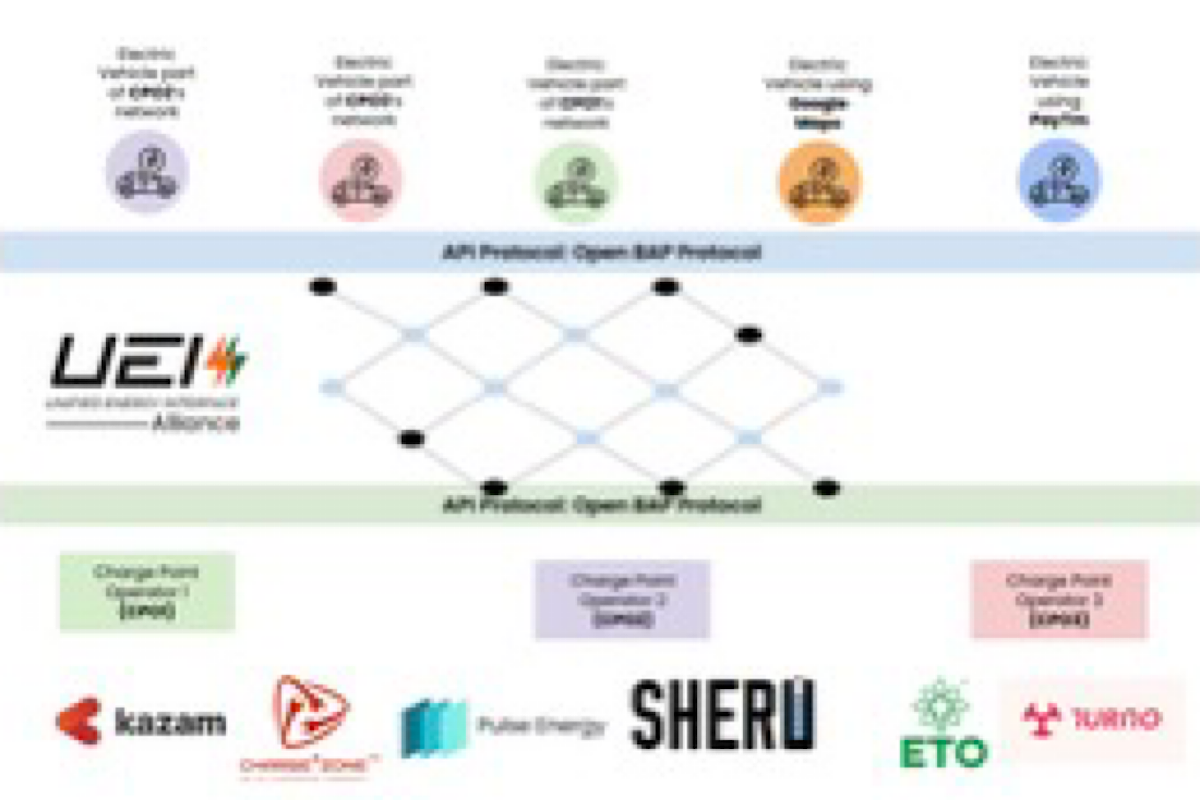
UEI is the recommended interoperability standard as per the Department of Science and Technology.

Chip maker Intel on Friday said that it appointed Santhosh Viswanathan to head the India region business as its Managing Director (MD).

To judge the battery life of any computer, measuring real-world performance like browsing the web, word processing, multimedia editing, etc. is a tough one.




