The ability to grow and reproduce is a fundamental property of living organisms. Whether an organism is composed of a single cell or trillions of cells, individual cells must be able to grow and divide in an appropriately regulated fashion.
Cell growth is accomplished through the synthesis of new molecules of proteins, nucleic acids, carbohydrates and lipids. As the accumulation of these molecules causes the volume of a cell to increase, the plasma membrane grows to prevent the cell from burst-ing.
But cells cannot continue to enlarge indefinitely; as a cell grows larger, there is an accompanying decrease in its surface area/volume ratio and hence in its capacity for ef-fective exchange with the environment.
Advertisement
Therefore, cell growth is generally accompanied by cell division, whereby one cell gives rise to two new daughter cells (The term daughter is used by convention and does not indicate that cells have gender).
For single-celled organisms, cell division increases the total number of individuals in a population. In multicellular organisms, cell division either increases the number of cells, leading to growth of the or-ganism, or replaces cells that have died. In an adult human, for example, about two million stem cells in bone marrow divide every second to maintain a constant num-ber of red blood cells in the body. When cells grow and divide, the newly formed daugh-ter cells are usually genetic duplicates of the parent cell, containing the same (or virtually the same) DNA sequences.
Therefore, all the genetic information in the nucleus of the parent cell must be duplicated and carefully distributed to the daughter cells during the division process.
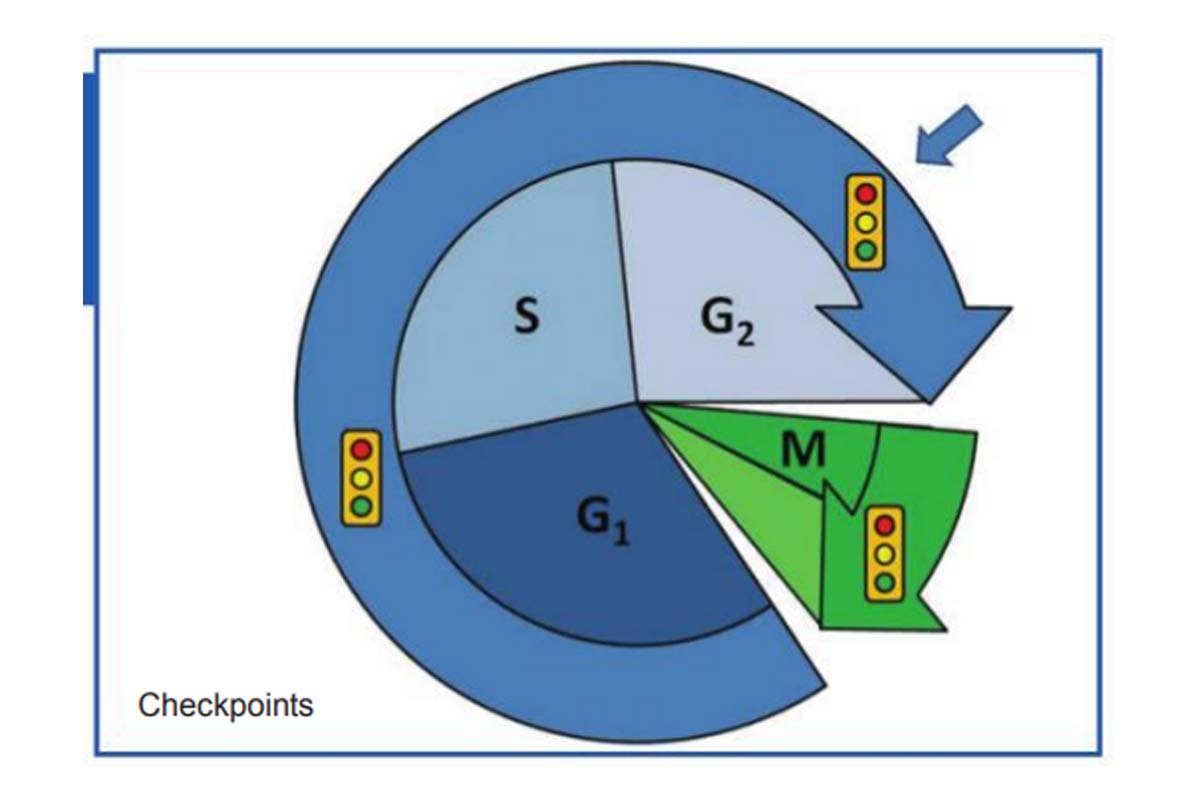
In accomplishing this task, a cell passes through a series of discrete stages, collectively known as the cell cycle. The cell cycle begins when two new cells are formed by the division of a single parental cell and ends when one of those cells divides again into two cells.
To early cell biologists studying eukaryotic cells with the mi-croscope, the most dramatic events in the life of a cell were those associated with the point in the cycle when the cell actually divides. This division process, called the M phase, involves two overlapping events in which the nucleus divides first and the cytoplasm second. Nuclear division is called mitosis, and the division of the cytoplasm to pro-duce two daughter cells is termed cytokinesis.
The stars of the mitotic drama are the chromosomes.
The beginning of mitosis is marked by condensation (coiling and folding) of the cell’s chromatin, which generates chromosomes that are thick enough to be individually discernible under the micro-scope.
Because DNA replication has already taken place, each chromosome actually consists of two chromosome copies that remain attached to each other until the cell divides. As long as they remain attached, the two new chromosomes are referred to as sister chromatids.
As the chromatids become visible, the nuclear envelope breaks into fragments. Then, in a stately ballet guided by the mi-crotubules of the mitotic spindle, the sister chromatids separate and — each now a full-fledged chromosome — move to opposite ends of the cell.
By this time, cytokinesis has usually begun, and new nuclear membranes envelop the two groups of daughter chromosomes as cell division is completed. While visually striking, the events of the mitotic phase account for a relatively small portion of the total cell cycle; for a typical mammalian cell, the mitotic phase usually lasts less than an hour.
Cells spend the majority of their time in the growth phase between divisions, called interphase. Most cellular contents are synthesised continuously during interphase, so cell mass gradually increases as the cell approaches division.
During interphase the amount of nuclear DNA doubles, and experiments using radioactive DNA precursors have shown that the new DNA is synthesised during a particular portion of interphase named the S phase (S for synthesis). A time gap called G1 phase separates the S phase from the preceding M phase, and a second gap, the G2 phase, separates the end of S phase from the beginning of the next M phase.
Although the cells of a multicellular organism divide at varying rates, most studies of the cell cycle involve cells growing in culture, where the length of the cycle tends to be similar for different cell types. We can easily determine the overall length of the cell cycle — the generation time — for cultured cells by counting the cells under a microscope and determining how long it takes for the population to double.
In cultured mammalian cells, for example, the total cycle usually takes about 18-24 hours. Once we know the total length of the cycle, it is possible to determine the length of specific phases. To determine the length of the S phase, we can expose cells to a radioactively labelled DNA precursor for a short period of time and then examine the cells by autoradiography.
The fraction of cells with silver grains over their nuclei represents the fraction of cells that were somewhere in S phase when the radioactive compound was available. When we mul-tiply this fraction by the total length of the cell cycle, the result is an estimate of the average length of the S phase.
For mammalian cells in culture, this fraction is often around 0.33, which indicates that S phase is about six to eight hours in length. Similarly, we can estimate the length of the M phase by multiplying the generation time by the percentage of the cells that are actually in mitosis at any given time.
This percentage is called the mitotic index.
The mitotic index for cultured mammalian cells is often about three to five per cent, which means that M phase lasts less than an hour (usually 30-45 minutes). In contrast to the S and M phases, whose lengths tend to be quite similar for different mammalian cells, the length of G1 is quite variable, depending on the cell type. Although a typical G1 phase lasts 8-10 hours, some cells spend only a few minutes or hours in Gl, whereas others are delayed for long periods of time. During Gl, a major “decision” is made as to whether and when the cell is to divide again. Cells that become arrested in Gl, awaiting for a signal that will trigger re-entry into the cell cycle and a commitment to divide, are said to be in G0 (G zero).
Other cells exit from the cell cycle entirely and undergo terminal differentiation, which means that they are destined never to divide again; most of the nerve cells in our body are in this state. In some cells, a transient arrest of the cell cycle can also occur in G2. In general, however, G2 is shorter than Gl and is more uniform in duration among different cell types, usually lasting 4-6 hours.
The writer is associate professor and head, department of botany, Ananda Mohan College, Kolkata.
Advertisement









